* 2005/07/27 - 2005/07/28
요즘에는 연구실 동기가 쓰고 있는 논문을 같이 고치고 있다. 처음에는 그냥 조금씩 코멘트만 하다가 지금은 더 깊이 개입하여 전반적인 내용, 논리전개 등도 함께 얘기하고 있다. 잡음이라고 하지 않고 노이즈라고 한 것은 노이즈가 꼭 잡'음'만 가리키는 것이 아니기 때문이다. 그럼에도 일반적으로 잡음이라고 번역한다.
우선 다루고 있는 대상은 동물의 신경세포, 즉 뉴런이다. 뉴런도 세포이므로 세포막이 있는데 이 세포막 안과 밖의 이온들(전하를 띤 입자, 여기서는 주로 칼륨, 나트륨, 칼슘 등이 있다)에 의해 전위차가 생긴다. 세포막에는 특정 이온의 통로(channel) 역할을 하는 단백질이 있어서 이것이 특정 이온의 농도를 조절한다. 뉴런 안쪽에는 칼륨이온의 농도가 높고 바깥쪽에는 나트륨이온의 농도가 높다.
뉴런은 생체 안의 신호를 전달하는데 펄스 형태의 전기 신호가 생기기도 하고 흐르기도 한다. 이러한 펄스를 활동전위라고 한다. 뉴런 안쪽의 전위는 평소에 약 -70mV를 유지하고 있는데 이를 휴지 전위(resting potential)라고 한다. 어떤 자극에 의해 나트륨이온통로가 열리고 세포막 바깥의 나트륨이온이 세포 안으로 들어오면서 전위가 높아지다가 어떤 문턱값(-55mV)에 이르면 탈분극화(depolarization)가 일어난다. 그러면 전위가 계속 높아지고 펄스가 시작된다(triggered). 나트륨이온통로는 수 ms(milisecond, 즉 0.001초)만에 닫히고 바로 칼륨이온통로가 열리면서 뉴런 안의 칼륨이온들이 바깥으로 나간다. 그러면서 전위가 다시 낮아진다. 위에 링크되어 있는 페이지에 이 과정이 설명되어 있다.가 아니라 실은 저 페이지의 내용을 거의 번역하다시피 하고 있는 중이다. 이렇게 전위가 높아졌다가 낮아지는 것을 활동전위라 부르고 수 ms의 활동전위 이후에 회복기(refractory period)가 이어진다.
이렇게 만들어진 펄스는 그냥 생겼다 없어지는 것이 아니라 뉴런의 축색돌기(axon)를 따라 전파된다. 한 뉴런의 축색돌기와 다른 뉴런의 수상돌기(dendrite)가 만나는 곳을 시냅스라고 하는데 시냅스를 통해 이러한 신호가 전달된다. 신호전달은 화학물질에 의한 방법과 전기신호가 직접 전달되는 방법이 있다.
1952년 Hodgkin과 Huxley는 뉴런의 활동전위를 미분방정식으로 모형화한 연구를 발표한다. 또한 이들은 오징어의 커다란 뉴런으로부터 직접 전위, 전류 등을 측정했는데 이들은 이 연구로 노벨상을 수상하기도 했다. 이들은 뉴런을 축전기와 가변저항이 있는 전기회로로 모형화했으며 이로부터 미분방정식을 만들어냈다. 이를 Hodgkin-Huxley 신경모형이라고 한다. 그런데 방정식이 복잡하여 컴퓨터로 풀지 않으면 풀기가 어렵다.
1960년대에 Fitz Hugh와 Nagumo 등은 더 간단하면서도 활동전위의 모양을 잘 재현해내는 모형을 제시한다. 나도 논문 몇 개만 본 것이 전부여서 더 말은 못하겠고 이 모형을 더 단순화하여 해석적인 접근을 시도한 논문이 1997년에 <피지컬 리뷰 레터스(PRL)>에 실렸다.
연구실 동기의 석사논문을 참고하면, 신경모형에 노이즈를 고려한 연구는 1993년에 처음 이루어졌다. 노이즈를 고려한 신경모형이 무엇을 의미하는지를 살펴보자.
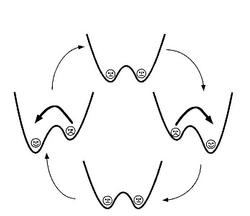
http://www.physik.uni-bielefeld.de/theory/cm/research/projects/SR.html
용수철 끝에 매달린 물체가 진동하는 것도, 실에 매달린 진자가 진동하는 것도 일반적으로 떨개(oscillator)라고 하는데 위의 시스템도 이중우물 + 약한 주기적인 힘 + 적당한 노이즈가 작용하는 떨개라고 볼 수 있다. 노이즈가 너무 작으면 도움이 안되고, 너무 크면 시스템 자체가 제멋대로 움직이므로 늘 적당한 크기의 노이즈가 필요하며 이때 SR을 관찰할 수 있다.
이러한 떨개들이 상호작용하지 않는다면 떨개가 여러개가 있든 하나만 있든 달라지는 것은 없을 것이다. 하지만 이들이 서로 상호작용하도록 만들면 어떻게 될까? 즉 위의 떨개들을 일렬로 주욱 늘어놓고 각각의 떨개는 양 옆의 두 떨개로부터 영향을 받도록 한다. 양쪽의 위치의 평균보다 자신이 큰 값을 갖고 있다면 자신의 값을 낮추고, 그 반대라면 자신의 값을 높인다. 이렇게 하면 SR의 전체적인 효과가 더욱 커진다. 이렇게 배열로 인해 SR이 더 좋아졌다고 해서 이를 array enhanced SR, 즉 AESR이라고 부른다.
SR과는 조금 다른 것으로 결맞는 공명(coherence resonance; CR)도 있다. 여기서는 외부의 주기적인 자극이 없이도 노이즈에 의해 공명 효과가 나타난다. 이를 모형화한 것 중의 하나가 앞 글에서 말한 Fitz Hugh-Nagumo(FHN) 모형이다. 여기서 '모형'이라는 것은 특정한 미분방정식을 뜻한다. 모든 모형이 미분방정식으로 나타나는 것은 아니다. 그냥 개념뿐일 수도 있고 규칙일 수도 있고 방정식일 수도 있다.
ε dx/dt = x - x^3 /3 - y
dy/dt = x + a + Dξ(t)
너무 수식이 없는 것 같아서 FHN 모형을 써봤다. dx/dt는 x를 t로 미분한다는 말이고 Dξ(t)는 노이즈를 뜻한다. ε은 1보다 아주 작은 값이어야 하고 a는 1보다 조금 크거나 -1보다 조금 작은 값이면 된다. 이때 노이즈의 세기 D를 조절해가면서 이 시스템이 어떻게 행동하는가를 보면 된다. 역시 적절한 크기의 D에서 CR이 가장 잘 나타난다는 것을 알 수 있다.
이것도 하나의 떨개인데, 여러개의 떨개가 상호작용하게 하면 AESR과 비슷한 결과를 얻을 수 있다. 이를 AECR이라고 한다. 상호작용에는 세 가지 요소가 있다. 상호작용의 방법, 상호작용의 세기, 연결구조다. 방법 중의 하나가 앞에서 소개한 것이다. 즉 양 옆의 x의 평균을 쫓아가기. 상호작용의 세기는 얼마나 빨리 쫓아가느냐이며, 연결구조는 양 옆만 볼 것인지 앞뒤도 볼 것인지 아니면 멀리 있는 뉴런까지 볼 것인지에 관한 얘기다. 다른 조건이 다 똑같아도 연결구조에 따라 다른 결과가 나올 수 있다.
흐름을 정리하면, 뉴런을 떨개로 모형화할 수 있고, 떨개에 적당한 노이즈를 가하여 공명이 일어나게 할 수 있으며(SR 또는 CR), 실제로 뉴런에 노이즈가 가해지는 것을 SR/CR로 모형화하여 이해할 수 있다. 여러개의 뉴런이 상호작용하는 시스템은 AESR/AECR 등으로 이해할 수 있다.
위의 FHN 모형을 다시 보자. FHN에서는 보통 ε=0.01, a=1.05 정도로 놓는다. 노이즈가 없는 경우, 즉 D = 0 일 때를 보자. a에 대해서만 설명을 하면, a가 1보다 크거나 -1보다 작은 값으로 주어질 때 x, y는 처음 어떤 값으로부터 시작을 하든 특정한 x, y 값(즉, x = -a, y = a^3 /3 - a)으로 빨려들어간다. 그리고 그곳에 고정되는데 xy 평면 위에서 보자면 어떤 점에 고정되어 버린다. 그래서 이를 '고정점'이라고 부른다. 그런데 a가 1과 -1 사이의 값으로 주어지면 초기 조건이 어떻든간에 xy 평면 위에서 특정한 폐곡선 위를 뱅뱅 돈다. 이를 '한계순환(또는 끝돌이; limit cycle)'이라고 한다.
CR 현상을 보기 위해서는 a를 1보다 야악간 크거나 -1보다 야악간 작은 값으로 주고, 노이즈 즉 D를 적당히 주면 된다. a가 1보다 크거나 -1보다 작으면 고정점에 붙어버리는데, 노이즈에 의해 계속 고정점으로부터 밀려났다가 다시 고정점으로 끌려오기를 되풀이한다. 그런데 노이즈가 조금 더 커지면 a가 1과 -1 사이일 때의 폐곡선을 따라 한바퀴 뱅 돌아서 다시 고정점으로 되돌아온다. 뉴런으로 보자면, 고정점 근처에서 지글지글 거리는 것이 휴지 전위에 해당하고, 앞서의 폐곡선을 따라 뱅 돌아오는 것이 활동전위에 해당한다.
그건 그렇고, 왜 여러 뉴런들이 상호작용하도록 해주면 더 공명이 잘 일어나는 것일까? 각 뉴런은 자신과 연결된 뉴런들의 x값들을 평균내어 자신의 x와 비교하여 이웃뉴런들의 평균을 쫓아가려고 한다. 뉴런이 활동전위를 겪는 것을 발화(firing)한다고도 하는데 노이즈에 의해 발화하기도 하고, 노이즈가 적어도 이렇게 이웃들에 의해 발화하기도 한다. 이렇게 뉴런들이 서로서로 발화하는 것을 도와주는데 이를 상호흥분(원래 mutual excitation인데, 맞는 번역인가?)이라고 한다. 노이즈가 도와주다못해 이웃들까지 도와주므로 공명이 더 잘 일어날 수 있을 것이다.라고 대충 이해해볼 수 있다.
노이즈의 긍정적 효과는 앞서 소개한 시스템뿐만 아니라 좀더 많은 현상에 적용해볼 수도 있다. 특히 사회현상에 적용해보는 것은 어떨까? 꼭 신경모형이 아니더라도 이미 노이즈에 대해, 그 적용에 대해 많은 연구가 이루어지고 있다. 생명체의 변이도 실은 노이즈에 의한 현상이 아닌가. 그렇다면 진화가 가능한 것도, 진화 그 자체도 노이즈에 의한 무엇이 될 수 있을 것이다.
앞의 노이즈 이야기에서 다룬 내용들에 관한 논문들을 여기에 소개한다.
A. C. Scott, "The electrophysics of a nerve fiber", Reviews of Modern Physics 47, 487 (1975)
- <신경섬유의 전기물리학>이라는 이 논문은 당시까지 신경모형을 주욱 정리해놓은 것인데, 49쪽짜리다. 상세하고 길어서 일부분만 보았다.
J. F. Lindner et al., "Array Enhanced Stochastic Resonance and Spatiotemporal Synchronization", Physical Review Letters 75, 3 (1995)
- 1차원 배열에 의한 SR을 연구한 논문.
Z. Gao, B. Hu and G. Hu, "Stochastic resonance of small-world networks", Physical Review E 65, 016209 (2001)
- 좁은 세상 연결망으로 연결된 신경모형에서의 SR을 연구한 논문.
A. S. Pikovsky and J. Kurths, "Coherence Resonance in a Noise-Driven Excitable System", Physical Review Letters 78, 775 (1997)
- FHN 모형을 위한 간단한 해석적 접근을 시도한 논문.
B. Hu and C. Zhou, "Phase synchronizatoin in coupled nonidentical excitable systems and array-enhanced coherence resonance", Physical Review E 61, R1001 (2000)
- 1차원 배열에서 FHN의 CR을 뉴런마다 다른 변수를 도입하여 연구한 논문.
C. Zhou, J. Kurths and B. Hu, "Array-Enhanced Coherence Resonance: Nontrivial Effects of Heterogeneity and Spatial Independence of Noise", Physical Review Letters 87, 098101 (2001)
- 역시 1차원 배열에서 FHN의 CR을 연구한 것인데 노이즈의 국소성(locality)을 변화시키면서 그 효과를 연구한 논문.